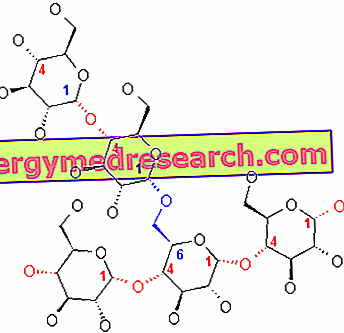
Glikogenas yra α-gliukozės makromolekulė (apie 400 milijonų daltonų molekulinė masė), kurioje yra daugiausia α-1, 4 glikozidinių ryšių ir santykių 1:10 santykis dėl α-1, 6 glikozidinių ryšių.
Glikogenas yra atsarginė medžiaga, kuri nuolat skaidoma ir rekonstituojama; viso kūno ląstelių masėje yra apie 100 g glikogeno: dauguma jų yra kepenyse, kur jis yra mobilus ir todėl gali būti naudojamas kaip rezervas kitiems organams (glikogenas raumenyse nėra mobilus).
Visi fermentai, skatinantys glikogeno degradaciją ir sintezę, yra citoplazmoje, todėl mums reikia koregavimo sistemos, kuri padėtų vieną neaktyvų neaktyvų, kai kitas yra aktyvus: jei yra gliukozės, pastaroji paverčiama glikogenu (anabolizmas), kuris yra rezervas, atvirkščiai, jei yra gliukozės paklausa, tada glikogenas degraduojamas (katabolizmas).
Fermentas, daugiausia susijęs su glikogeno skaidymu, yra glikogeno fosforilazė ; šis fermentas gali suskaidyti α-1, 4 glikozidinę jungtį, naudodamas kaip lytinį agentą neorganinį ortofosfatą: skaidymas vyksta fosforolitiniu būdu ir gaunamas gliukozės 1-fosfatas.
Penkiais arba šešiais vienetais iš šakotojo taško glikogeno fosforilazės fermentas nebegali veikti, todėl jis išsiskiria nuo glikogeno ir pakeičiamas deramifikuojančiu fermentu, kuris yra transferazė : katalizinėje šio fermento vietoje yra histidinas, kuris leidžia perkelti tris sacharido vienetus į artimiausią glikozidų grandinę (histidinas atakuoja pirmąją gliukozės molekulės anglies). Minėtas fermentas yra glikoziltransferazė ; pasibaigus šio fermento veikimui, tik vienas gliukozės blokas lieka su pirmuoju anglies junginiu prie pagrindinės grandinės gliukozės šeštosios anglies. Paskutinis šoninės grandinės gliukozės vienetas išsiskiria fermento α-1, 6 glikozidazės poveikiu (šis fermentas sudaro antrą fermento dalį, kuri yra nereikšminga); kadangi glikogeno pasiskirstymas yra santykiu 1:10, nuo visiško makromolekulės skilimo gauname apie 90% gliukozės 1-fosfato ir apie 10% gliukozės.
Minėtų fermentų poveikis leidžia pašalinti šalutinę grandinę iš glikogeno molekulės; šių fermentų aktyvumas gali būti kartojamas, kol atsiras visas grandinės degradavimas.
Apsvarstykite hepatocitus; gliukozė (įsisavinama per mitybą), kai ji patenka į ląstelę, paverčiama 6-fosfato gliukoze ir taip aktyvuojama. Gliukozės 6-fosfatas, dėl fosfoglukomutazės poveikio, yra transformuojamas į gliukozės 1-fosfatą: pastarasis yra tiesioginis biosintezės pirmtakas; biosintezėje naudojama aktyvuota cukraus forma, kurią sudaro cukrus, prijungtas prie difosfato: paprastai uridildifosfatas (UDP). Tada 1-fosfato gliukozė paverčiama UDP-gliukoze; šis metabolitas patenka į glikogeno sintezės veikimą, kuris gali surišti UDP-gliukozę prie ne mažinančio augančio glikogeno galo: gaunamas pailgintas glikozido vieneto ir UDP glikogenas. UDP fermentą nucleosidedifosfochinasi paverčia UTP, kuris grįžta į cirkuliaciją.
Glikogeno degradacija atsiranda dėl glikogeno fosforilazės, kuris išskiria gliukozės molekulę, ir paverčia jį gliukozės 1-fosfatu. Vėliau fosfoglukomutazė konvertuoja 1-fosfato gliukozę į 6-fosfato gliukozę.
Glikogenas sintetinamas, ypač kepenyse ir raumenyse: organizme yra 1–1, 2 hektolitrų glikogeno, išplatintoje per visą raumenų masę.
Miocitų glikogenas yra energijos rezervas tik šiai ląstelei, o kepenyse esantis glikogenas taip pat yra rezervas kitiems audiniams, ty jis gali būti siunčiamas, kaip gliukozė, kitoms ląstelėms.
Tada gliukozės 6-fosfatas, gautas raumenyse iš glikogeno degradacijos, tada energijos poreikio atveju siunčiamas į glikolizę; kepenyse 6-fosfatas gliukozės pavidalu paverčiamas gliukozės 6-fosfato fosfataze (hepatocitams būdingas fermentas) ir nukreipiamas į kraujotakos srautą.
Glikogeno sintezė ir glikogeno fosforilazė veikia tiek nesumažinančiais glikogeno vienetais, todėl turi būti hormoninis signalas, kuris komanduoja vieną kelią ir kitą bloką (arba atvirkščiai).
Laboratorijoje buvo galima pailginti glikogeno grandinę, panaudojant glikogeno fosforilazę ir naudojant labai didelę koncentraciją gliukozės 1-fosfatu.
Ląstelėse glikogeno fosforilazė katalizuoja tik degradacijos reakciją, nes metabolitų koncentracijos yra tokios, kad perkelia toliau nurodytos reakcijos pusiausvyrą į dešinę (t.y., glikogeno degradacijai):

Mes matome glikogeno fosforilazės veikimo mechanizmą: acetalinis deguonis (kuris veikia kaip tiltas tarp gliukozės vienetų) jungiasi su fosforilo vandeniliu: tarpinė reakcija susidaro karbokarte (gliukozėje, kuri yra galūnės), prie kurių labai greitai susieja fosforilą (Pi).
Glikogeno fosforilazei reikia kofaktoriaus, kuris yra piridokso fosfatas (ši molekulė taip pat yra transaminazių kofaktorius): jis tik iš dalies fosforilavo protoną (piridokso fosfatą supa hidrofobinė aplinka, kuri pateisina protonų, prie kurių prisijungta, buvimą), Fosforilas (Pi) gliukogenui gali duoti protoną, nes šis fosforilas vėl atgimsta, protonas iš iš dalies apsaugoto piridokso fosfato fosforilo. Tikimybė, kad esant fiziologiniam pH fosforilui neteks protono ir lieka visiškai deprotonuota, yra labai maža.
Dabar pažiūrėkime, kaip veikia fosfoglukomutazė. Šis fermentas katalizinėje vietoje turi fosforilintą serino liekaną; serinas sukelia fosforilą 1-fosfatui (šeštoje padėtyje): trumpą laiką gliukozė susidaro 1, 6-bisfosfatas, tada serinas yra refrofosfiliuojamas, fosforilą perimant pirmoje padėtyje. Fosfoglukuso mutazė gali dirbti abiem kryptimis, ty 1-fosfato gliukozės konversija į 6-fosfato gliukozę arba atvirkščiai; jei gaminama 6-fosfato gliukozė, ji gali būti tiesiogiai siunčiama į glikolizę, į raumenis arba transformuojama į gliukozę kepenyse.
Uridilo fosfoglukuso transferazės (arba UDP gliukozės pirofosforilazės) fermentas katalizuoja gliukozės-1-fosfato perkėlimo reakciją į UTP, atakuodamas fosforilą a.
Pirmiau aprašytas fermentas yra pirofosforilazė: šis pavadinimas atsirado dėl to, kad priešinga reakcija, kaip aprašyta, yra pirofosforilinimas.
UDP gliukozė, gauta kaip aprašyta, gali pailginti monosacharido vieneto glikogeno grandinę.
Galima išsivystyti reakciją į UDP gliukozės susidarymą pašalinant produktą, kuris yra pirofosfatas; pirofosfatazės fermentas pirofosfatą paverčia dviem ortofosfato molekulėmis (anhidrido hidrolizė) ir tokiu būdu išlaiko pirofosfato koncentraciją taip mažai, kad UDP gliukozės susidarymo procesas yra termodinamiškai skatinamas.
Kaip minėta, glikogeno sintezės dėka UDP gliukozė gali pailginti glikogeno grandinę.
Poveikiai (santykiu 1:10) yra susiję su tuo, kad, kai glikogeno grandinė susideda iš 20-25 vienetų, įsijungia šakotasis fermentas (kuriame yra katalizatoriaus vietoje esantis istidinas), galintis perkelti 7 serijų seriją. -8 glikozidiniai vienetai pasroviui nuo 5-6 vienetų: taip sukuriamas naujas filialas.
Nervų klausimu arba, jei dėl fizinės jėgos reikia energijos, adrenalinas išsiskiria nuo antinksčių.
Adrenalino (ir norepinefrino) tikslinės ląstelės yra kepenų, raumenų ir riebalinio audinio ląstelės (pastarojoje yra trigliceridų skaidymas ir riebalų rūgščių cirkuliacija: todėl mitochondrijose gaminama gliukozė 6 -fosfatas, siunčiamas į glikolizę, o adipocituose gliukozės 6-fosfatas transformuojamas į gliukozę 6-fosfato fosfatazės fermentu ir eksportuojamas į audinius).
Pažiūrėkime, dabar yra adrenalino veikimo būdai. Adrenalinas jungiasi prie receptoriaus, esančio ląstelių membranoje (miocitų ir hepatocitų), ir tai lemia signalo vertimą iš išorės į ląstelės vidų. Aktyvuojama baltymų kinazė, kuri tuo pačiu metu veikia sistemose, reguliuojančiose glikogeno sintezę ir skaidymą:
Glikogeno sintezė egzistuoja dviem formomis: defosforilinta (aktyvi) forma ir fosforilinta (neaktyvi) forma; baltymų kinazė fosforilina glikogeno sintezę ir blokuoja jo veikimą.
Glikogeno fosforilazė gali egzistuoti dviem būdais: aktyvi forma, kurioje yra fosforilintas serinas ir neveikli forma, kurioje serinas yra defosforilintas. Glikogeno fosforilazę gali aktyvuoti glikogeno fosforilazės kinazė . Glikogeno fosforilazės kinazė yra aktyvi, jei ji yra fosforilinta ir neaktyvi, jei ji yra defosforilinta; baltymų kinazės substratas yra glikogeno fosforilazės kinazė, ty ji yra pajėgi fosforilinti (ir todėl aktyvuoti) pastarąją, kuri savo ruožtu suaktyvina glikogeno fosforilazę.
Baigus adrenalino signalą, jis taip pat turi nutraukti savo poveikį ląstelėje: tada fosfatazės fermentai turi įtakos baltymų rūšims.